


Билатеральные комплексы бочонковых колонок первичной соматосенсорной коры мозга являются модулями восприятия пространства
В.Н. Ласков
НИИ нейрокибернетики им. А.Б.Когана, РГУ, Россия
Исследованы пространственные параметры рецептивных полей (РП) нейронов в корковом представительстве С1 коры мозга крыс латеральных и медиальных рядов вибрисс. Показано, что в бочонковых колонках представительства латеральных вибрисс РП имеют овальную форму и ориентированы вдоль одноименного ряда вибрисс. На каждую колонку проецируются все вибриссы контралатеральной стороны. Размер, форма и структура РП зависят от амплитуды стимула и общего уровня возбуждения колонки. Колонки в представительстве медиальных вибрисс имеют билатеральные РП: их ипсилатеральное полуполе представлено медиальными рядами вибрисс с низкой проявляемостью реакций, а контралатеральное - не включает нижние ряды вибрисс. Исследованы реакции нейронов и паттерны фокальных вызванных потенциалов (ВП) билатеральных комплексов (БК) бочонковых колонок при разном пространственном положении тактильных стимулов. Показана специфичность паттернов импульсной и медленной суммарной активности в БК положению стимулов в рецептируемом пространстве. Показана повышенная чувствительность колонок представительства передних, задних, верхних и нижних вибрисс к наиболее часто реализуемым в поведении крыс направлениям их отклонения. Исследовано пространственное распределение в бочонковых колонках дирекционально избирательных нейронов и определены их сетевые свойства. Расположение дирекциональных миниколонок в пространстве бочонковой колонки соответствует радиальной симметрии рецептируемого пространства. Дирекциональная настройка нейронов на входе колонки обеспечивается топикой представительства рецепторов фолликула, а на ассоциативном и выходном уровнях - системой тормозных интернейронов.
Введение
Способность человека и животных определять местоположение стимулов в пространстве, в том числе траектории их движения и ориентацию, а также оценивать размер, форму, текстуру поверхности и многие другие признаки называется пространственным восприятием. Этой способностью наделены органы чувств высших позвоночных и беспозвоночных животных. Общим проявлением механизма восприятия пространства в жизнедеятельности человека и животных являются бинарные эффекты пространственного различения: бинокулярное зрение, бинауральный слух, дириническое обоняние и бимануальное (в широком смысле) осязание /1/. В эволюции механизмы дальномерности и стереосенсорности возникли на основе билатерального плана строения тела активно-подвижных животных, у которых для успешной ориентации в пространстве сформировались парные локомоторные и сенсорные органы. Они воспитали, по образному выражению, билатеральный мозг с его высшими отделами, парными большими полушариями, которые и являются базой механизма восприятия пространства.
К настоящему времени исследованы морфология и физиология межполушарных связей в составе мозолистого тела, которое обеспечивает нейронные механизмы бинарных эффектов в восприятии пространства /2,3,4/. В большей мере это касается механизмов стереопсиса и других известных феноменов зрительного восприятия /4/, а также пространственного слуха /5/. Несмотря на актуальность проблем, связанных с выяснением принципов нейронной организации систем, ответственных за билатеральное тактильное восприятие, а также способов кодирования пространственных параметров тактильных стимулов, разработаны они лишь в первом приближении.
Ранее нами получены результаты /6,7/, свидетельствующие, что каждая из бочонковых колонок в корковом представительстве вибрисс мозга крысы включает в свой состав миниколонки нейронов, каждая из которых детектирует одну из дирекциональностей движущегося тактильного стимула. Было высказано предположение, что схема расположения дирекциональных миниколонок в бочонковой колонке инвариантна радиальной симметрии возможных направлений сгибания вибриссы. Кодирование дирекциональности в корковом модуле топического представительства, согласно нашему представлению, осуществляется за счет диссимметризации его исходного сбалансированного уровня возбуждения, превращающей колонку в функциональный диполь с соответствующим дирекциональности вектором активности.
Кроме того, было показано, что на билатеральных гомотопных колонках в С1 коры мозга крыс конвергируют, по крайней мере, две топически организованные рецептивные поверхности: рецепторы вибрисс - с полным перекрестом афферентных путей и рецепторы, находящейся под вибриссами шерсти, имеющие двустороннее представительство в бочонковых колонках. Это создает предпосылки для кодирования местоположения и других пространственных параметров, осязаемых с помощью вибрисс и шерсти под ними тактильных стимулов, пространственными паттернами нейронного возбуждения в симметричной структуре БК бочонковых колонок /8/.
Целью данной работы явилось дальнейшее изучение обнаруженных нами особенностей нейронной организации ответственных за билатеральное тактильное восприятие БК бочонковых колонок в С1 коры мозга крыс, а также нейронных механизмов кодирования пространственных параметров тактильных стимулов.
Исходной гипотезой в данном исследовании явилось представление о том, что БК, как специализированные сенсорные единицы пространственного восприятия, являются функциональными диполями, градиенты активности которых задаются пространственными параметрами тактильных стимулов. При этом, предполагаемое нами, но не показанное в специальной литературе для данного сенсорного прибора, перекрытие РП нейронов в представительстве медиальных рядов вибрисс служит нейронной моделью средней линии тела, как это описано для зрительной системы /4/, и своеобразной осью координат при оценке местоположения семейства точек на поверхности объекта.
Для проверки этой гипотезы были исследованы размеры, форма и степень билатерального перекрытия РП нейронов бочонковых колонок в представительстве латеральных и медиальных вибрисс, а также паттерны импульсной активности нейронов и фокальных ВП на разных уровнях БК бочонковых колонок при разном пространственном положении тактильных стимулов различной дирекциональности.
Обзор литературы
Роль вибрисс в пространственной ориентации.
Крысы, наряду со многими другими млекопитающими, ведущими сумеречный образ жизни, ориентируются в пространстве с помощью вибрисс. У многих видов вибриссы двигаются, мерцают, и, чем тоньше пространственный анализ, тем короче и чаще ощупывающие взмахи вибрисс. Имеются данные о том, что оптимальная частота исследовательских движений вибрисс соответствует тетаритму в ЭЭГ-активности крыс /9/. С помощью вибрисс осуществляется дистантный анализ тактильной информации - они являются переходным этапом между рецепторными аппаратами контактного и дистантного типа. Длина вибрисс у некоторых видов животных сопоставима с длиной туловища: у песчанки вибриссы имеют длину 15 см при длине туловища 10 см. Особую роль вибриссы играют в ближней ориентации. С их помощью крысы получают информацию о неохваченном зрением пространстве, "мертвой зоне", которая образуется вследствие латерального расположения глаз и включает область перед мордой и над ней /10,11/.
Удаление вибрисс нарушает у крыс процессы направленной локомоции, поддержания равновесия, ориентации и манипуляции с предметами /12,13/, зрительно обусловленной оценки глубины пространства /13/. Как показывают наблюдения в "открытом поле", крысы получают с помощью вибрисс дополнительную информацию при манипуляции с незнакомыми предметами. При этом они обходят вокруг предмета, непрерывно ощупывая его с помощью вибрисс. И уже последующие манипуляции с предметами полностью зависят от результатов их предварительного осязания: животные перекатывали только объемные и относительно плотные предметы, переносили в зубах или грызли только мягкие и рыхлые предметы, поднимали зубами, не пытаясь воспользоваться передними лапками, совсем плоские, прилегающие к полу, предметы. Это свидетельствует о наличии в ходе ощупывания вибриссами процесса распознавания /14/. По-видимому, в процессе исследовательского поведения устанавливаются наиболее общие и значимые для животного признаки предметов и ближайшего окружающего пространства. Это - элементы формы и текстуры поверхности, наличие на пути преград или проходов, относительный размер которых требуется оценить, потоки воздуха с той или иной стороны, что имеет значение для успешного и адекватного поведения в условиях зрительного дефицита, обусловленного жизнью в затемненных местах.
У крыс вибриссы располагаются на верхней губе пятью рядами, которые ориентированы в каудо-ростральном направлении и условно обозначаются буквами А, В, С, D, Е, начиная сверху. В каждом ряду вибриссы нумеруют, начиная с задних крупных вибрисс. Уменьшение длины вибрисс в ростральном направлении от 30-50 мм до 4-5 мм у самого носа обеспечивает одновременный контакт их кончиков с осязаемым предметом при ощупывающих движениях в конце траекторий. Считают /15/, что это способствует более равномерной информационной загруженности отдельных волосков и связанных с ними вибриссковых нервов. Кроме того, упорядоченное расположение мелких и крупных вибрисс обеспечивает широкое разрешение тактильного анализа. Крупные вибриссы обеспечивают большую площадь рецептируемой поверхности, а густо расположенные мелкие вибриссы - высокую разрешающую способность. Билатеральные вибриссы верхних рядов (А и В) при исследовательских движениях осязают пространство, находящееся в непосредственной близости к средней линии тела. Они ощупывают с двух сторон предметы, занимающие правое и левое полуполя рецептируемого пространства. Мы назвали эти два ряда вибрисс медиальными, поскольку они в функциональном плане аналогичны кожным РП, переходящим через среднюю линию тела. Вибриссы рядов С, D и Е ориентированы в глубину своего полуполя, поэтому мы называем их латеральными. По-видимому, разная ориентация рядов вибрисс определяется качественными различиями воспринимаемой ими пространственной информации. Медиальные вибриссы имеют большее отношение к восприятию билатерально расположенных объектов, а латеральные - наряду с этим оценивают латерализацию объектов, их расстояние от средней линии тела в своем полуполе.
Межполушарные связи ПМСБ С1 коры мозга крыс.
Наибольшей плотностью межполушарных связей характеризуются корковые зоны представительства осевых частей сенсорной поверхности: вертикальный меридиан зрительных РП, а также те участки соматосенсорной коры, которые активируются кожными и суставными рецепторами, расположенными вблизи средней линии тела на туловище и голове, но не связывает корковые проекции конечностей /3,4/. Мозолистое тело связывает корковые нейроны таким образом, что их РП заходят вправо и влево от медианы тела, соединяя тем самым две половины рецептируемого пространства в единую панораму.
Организация каллозальных проекций постеромедиальной субзоны бочонков (ПМСБ) С1 коры мозга крыс и мышей детально исследована с помощью авторадиографических, транспортных и антероградных методов /16,17,18,19 и др./. Показано, что каллозальные волокна входят в субзону колончатыми пучками, которые в ряде случаев сливаются в более крупные пучки. Они поднимаются по краю или между бочонками, не контактируя с клетками проекционных модулей. Наибольшая плотность терминалей комиссуральных волокон наблюдается в слоях 1 и 3, а также в верхней части слоя 5 и в глубине слоя 6. Слоистое распределение терминальных разветвлений пучков каллозальных волокон соответствует дендритной организации пирамид слоя 3 и слоя 5. Эти пирамидные нейроны имеют густо ветвящиеся дендриты: базальные - в пределах того же слоя и апикальные - в верхней части слоя 3 и в слое 1, с которыми и контактируют терминали каллозальных афферентов.
Топография каллозальных проекций в ПМСБ характеризуется билатеральной адресностью. Нейроны, дающие начало комиссуральным проекциям в субзону противоположного полушария, одновременно являются постсинаптическими для таких же проекций из гомотопных пунктов С1. Их перикарионы сгруппированы в колонки, а последние располагаются между бочонками слоя 4, а также над и под бочонками во 2-3 слоях и в 5-6 слоях /19/. Гистологически не определено, получают ли таламическую афферентацию нейроны каллозальных колонок. Согласно /20/, эти колонки не активируются соматическими сенсорными стимулами; возможно, что у крысы имеет место сегрегация таламического и каллозального входов в данной субзоне. Не похоже, однако, что такое разделение является абсолютным для терминальных разветвлений пучков комиссуральных волокон. Их терминали распространяются очень широко в супрабочонковых слоях и входят в соприкосновение с дендритами клеток бочонка.
Большинство авторов /16,17,18,19/ отмечают, что корковое представительство вибрисс с С1 коры мозга крыс характеризуется достаточно слабым развитием каллозальных связей по сравнению с другими медианными областями проекции тела. Малое количество комиссуральных связей в представительстве вибрисс является, по мнению авторов, несколько неожиданным, т.к. представительство тройничного нерва у других видов имеет более тесную связь. Поэтому, из-за малой плотности каллозальных связей, многие авторы, склонны рассматривать вибриссы как добавочные сенсорные органы /21/, сравнимые с конечностями, корковые представительства которых лишены каллозальных входов.
Подробно ознакомившись с методикой этих гистологических исследований, мы обратили внимание на то, что изложенные в них результаты могут быть отнесены лишь к центральным бочонковым колонкам, которые являются проекцией латеральных вибрисс рядов С, D, Е. Бочонки в представительстве медиальных вибрисс рядов А и В, в определенном смысле, неудобны для гистологического и электрофизиологического исследований. Они располагаются более височно и мало доступны для введения микроэлектродов и перфузии красителей. Кривизна поля 2 (по Кригу, 1946) не позволяет получить на тангенциальных срезах коры мозга, сделанных на уровне слоя 4 (где отчетливо выделяются контуры отдельных бочонков), полную картину ПМСБ. Ее периферия, куда проецируются медиальные ряды, представлена на срезах не уплощенных препаратов более поверхностными слоями, где бочонковые колонки гистологически не выявляются. В предыдущем нашем исследовании /22/ в функционально идентифицированную бочонковую колонку ряда А вводили микродозы (0,12 мкл) примулина. Анализировались тангенциальные срезы соответствующих участков плаща коры мозга, приготовленные с резанием со стороны белого вещества /28/. На гистологических препаратах место введения флюорохрома было локальным, не выходящим за пределы слоя 5, и по диаметру не превышающим размеров бочонка, - 100-200 мкм. При просмотре замороженных срезов симметричных локусов коры обнаружены в верхней части слоя 5 плотные колонки светящихся нейронов пирамидного типа. Сопоставление этих результатов с результатами исследования, в котором выведение флюорохрома проводилось в бочонки ряда С, показало, что колонки в представительстве медиальных вибрисс ряда А содержат достоверно большее количество каллозальных нейронов по сравнению с бочонковыми колонками представительства латеральных рядов вибрисс.
В нейрофизиологических исследованиях /6,7,8/ показана организация в пределах бочонковых колонок С1 коры мозга крыс каллозальных входов от гомотопных и гетеротопных колонок противоположного полушария, а также степень перекрытия и характер взаимодействия в них каллозальных и таламических входов и выходов. Были показаны существенные различия пространственно-временной организации реакций каллозальных нейронов и распределения каллозальных афферентов в бочонковых колонках представительства медиальных и латеральных вибрисс. Так, в проекции медиальных вибрисс подавляющее большинство нейронов участвует в обработке каллозальной афферентации, тогда как в проекции латеральных вибрисс таких нейронов - лишь немногим более половины. Показана функциональная неоднородность структуры бочонковых колонок, выражающейся в разной степени перекрытия таламического и каллозального входов в составляющих ее миниколонках. По этому признаку в пределах бочонковой колонки выделены три разновидности миниколонок: с доминированием таламического входа, с доминированием каллозального входа и интегративные миниколонки - с равным весом обоих входов. При этом таламический и каллозальный входы были в определенном смысле парциальны: миниколонки, лишенные одного из входов, характеризовались более короткими ЛП на доминирующую афферентацию.
Обнаружены существенные различия также и в организации входов и выходов интегративных миниколонок. В миниколонках представительства латеральных вибрисс одни и те же нейроны являются входными для первичной и вторичной афферентации, а таламический выход изолирован от каллозальных влияний. В представительстве медиальных вибрисс таламический вход интегративной миниколонки изолирован от каллозального, но одни и те же нейроны являются выходными для таламической и каллозальной информации. Высказано предположение, что в различной функциональной организации бочонковых колонок представительства латеральных и медиальных вибрисс выражено разное качество обрабатываемой ими информации, касающейся, соответственно, степени латерализации тактильного стимула и его билатеральных пространственных свойств.
Кроме того, было показано, что гомотопные бочонковые колонки в представительстве медиальных вибрисс попарно адресно связаны двусторонними каллозальными афферентами и в силу этого представляют собой распределенную систему, названную нами билатеральным комплексом, который, по-видимому, является функциональной единицей более высокого уровня нейронной интеграции.
Методика экспериментов
Опыты выполнены на белых беспородных крысах обоего пола весом 120-200 г, оперированных под легким эфирным наркозом, а затем обездвиженных тубокурарином (1,5 мг/кг в/м), и переведенных на искусственное дыхание. Трахеотомия не производилась, воздух в легкие нагнетался через ноздри. Операционное поле подвергалось местной анестезии 0,5% раствором новокаина. Осуществлялась жесткая фиксация головы с помощью головодержателей. Череп трепанировался над ПМСБ корковой области С1 по координатам: 2-3 мм каудальнее брегмы и 5-6 мм латеральнее сагиттального шва. Импульсная активность корковых нейронов, а также фокальные ВП отводились стеклянными одиночными микроэлектродами, заполненными 2,5 М раствором NaCL, с сопротивлением около 5 мОм и вольфрамовыми в стеклянной изоляции полумикроэлектродами с сопротивлением 0,5-1 мОм, которые погружались в кору отвесно к ее поверхности. Контролем попадания электродов в ПМСБ служили фокальные ВП, предварительно прослушиваемые на аудиомониторе, которые возникали в ответ на легкое постукивание по вибриссам контралатеральной стороны мордочки крысы вначале вручную, а затем с помощью специальных тактильных датчиков.
Идентификация колонок. Сравнивая фокальные ВП, наблюдаемые на видеомониторе по их латентным периодам (ЛП), крутизне и амплитуде в ответ на сгибание вибрисс, устанавливали, как показано на рис.1,а, соответствие между бочонковой колонкой и центральной в РП. Для более точной идентификации центра РП нейронов производилась стимуляция ближайших к центру вибрисс со ступенчатым уменьшением амплитуды их отклонения (рис.1,б). Наблюдаемое при этом сужение РП до одной вибриссы в случае ее центрального положения имело место при наиболее низких амплитудах ее отклонения.
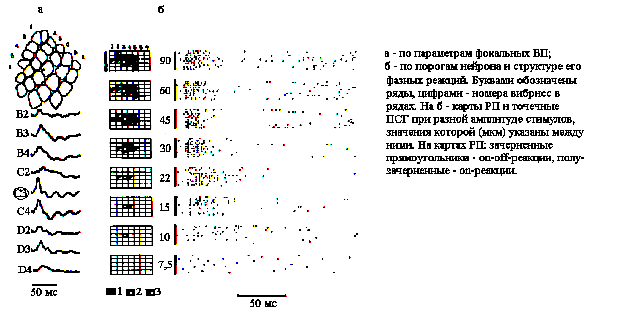
Рис. 1. Идентификация бочонковых колонок С1 коры мозга крысы.
Картирование РП.
Картирование РП нейронов производилось путем построения постстимульных гистограмм (ПСГ) реакций нейронов на последовательное отклонение вибрисс вначале контралатеральной, а затем ипсилатеральной стороны мордочки крысы. Амплитуда отклонения вибрисс при картировании превышала в десять раз ее пороговое значение, определяемое для центра РП.
Определение порога. При определении пороговых характеристик нейронов в качестве адекватного стимула использовано сгибание центральной в РП вибриссы с удержанием ее в отклоненном положении в течение 0,1-0,5 с. Чтобы избежать неконтролируемых смещений, более дальние от носа вибриссы, достигающие в длину 30-50 мм, укорачивались справа и слева до 5-10 мм. Механический датчик для определения пороговых характеристик нейронов представляет собой щуп, приклеенный на свободном конце пьезокерамической пластины, изготовленной в ОКБ "Пьезоприбор" РГУ, на которую подавалось напряжение от 60 до 1В с выхода электростимулятора ЭСЛ-2, задающего амплитуду отклонения на самом дальнем конце щупа в диапазоне от 90 до 1,5 мкм. Предварительная метрологическая проверка показала линейный характер зависимости между напряжением и амплитудой сгибания пьезокерамической пластины в используемом диапазоне. Поэтому непосредственно в опыте амплитуда отклонения щупа оценивалась по напряжению на выходе ЭСЛ-2. Серия стимулов определенной амплитуды предъявлялась ритмично с частотой 0,5 Гц. По 10-20 реализациям строились точечные и столбиковые ПСГ с бином 1-2 мс за период 100 или 500 мс. При определении пороговых характеристик нейронов рассматривались только on-эффекты стимуляции. Определялись параметры первичных компонентов возбудительных (В-) реакций на каждый из стимулов серии, а также усредненный паттерн разряда. Пороговым считалось такое значение амплитуды, при котором нейрон реагировал в серии повторных стимулов с вероятностью равной или большей 0,5.
Определение дирекциональности нейронов.
Для определения дирекциональной селективности нейронов бочонковых колонок использовался специальный тактильный датчик, представляющий собой блок четырех миниатюрных катушек индуктивности, отклоняющих щуп с приклеенной к нему вибриссой вверх, вниз, вперед и назад относительно мордочки животного. Восьмикоординатный тактильный датчик был выполнен компактно и состоял из четырех миниатюрных катушек индуктивности, сердечники которых выставлялись строго параллельно по углам квадрата со стороной 12 мм. В центре квадрата располагался щуп, укрепленный на площадке, на которой были зафиксированы сердечники катушек. Щуп проходил через центр тонкого стального диска, зафиксированного на щупе в непосредственной близости от плоскости, в которой находились сердечники катушек. При включении каждой из катушек диск притягивался к ее сердечнику и смещал щуп в соответствующем направлении. Одновременное включение двух соседних катушек отклоняло щуп в промежуточном направлении. Амплитуда отклонения по четырем взаимно перпендикулярным направлениям выравнивалась с помощью резисторов, регулирующих напряжение в каждой из катушек в отдельности. В начале опыта датчик устанавливался таким образом, чтобы его основные координаты обеспечивали отклонение вибриссы вверх-вниз, вперед-назад.
Дирекционально избирательными считались такие нейроны, первичные В-реакции которых на отклонение вибрисс по четырем взаимно перпендикулярным направлениям достоверно (Р<0,05) различались по ЛП, а по частоте следования импульсов давали двукратное различие первичных разрядов по двум противоположным либо перпендикулярным направлениям /23/.
Билатеральная стимуляция.
При исследовании паттернов импульсной и суммарной вызванной активности БК колонок стимуляция соответствующих вибрисс и шерсти под ними осуществлялась с помощью двух идентичных тактильных датчиков, изготовленных на основе реле типа РЭС 22. Вариация амплитуды углового отклонения вибрисс в этом случае осуществлялась за счет длительности подаваемых на датчики импульсов, а также за счет смещения щупа датчика вдоль вибриссы. Последняя процедура привлекательна тем, что, не меняя скорости и времени отклонения вибриссы, к которым чрезвычайно чувствительна данная система, но, помещая щуп датчика на все большем расстоянии от фолликула, удавалось очень точно задавать достаточно малые угловые отклонения вибриссы.
Импульсная активность и сопряженные с ней фокальные ВП регистрировались в диапазоне 2-2000 Гц, а усредняемые ВП - 0,3-30 Гц на входных фильтрах электроэнцефалографа ЧЭЭГ-03. Биоэлектрическая активность записывалась в опыте на магнитограф EАМ-500 и обрабатывалась по стандартным программам на ЭВМ IBM PC AT. Усреднение ВП производилось по 10-40 реализациям с квантом 1 мс за период 2 секунды.
Результаты исследования и их обсуждение
Размер и форма РП нейронов бочонковых колонок
Картированы В-зоны РП 82 нейронов разных уровней бочонковых колонок ПМСБ С1 коры мозга крыс. Исследования проводили в плане сравнения размеров и формы РП нейронов, находящихся в представительстве латеральных (преимущественно в бочонке С3) и медиальных (бочонки рядов В и А) вибрисс. В первой выборке нейронов (N=52), представляющей бочонок С3, размер РП при амплитуде стимула, равной десяти пороговым значениям, как это показано на рис.2а, варьировал от одной до 23 вибрисс. Около 11% нейронов, относящихся, преимущественно, к слою 4, имели одновибриссные РП. Амплитудные пороги у этих нейронов были наиболее низкими - от 3 до 15 мкм, при среднем значении 5,3+0,7 мкм.
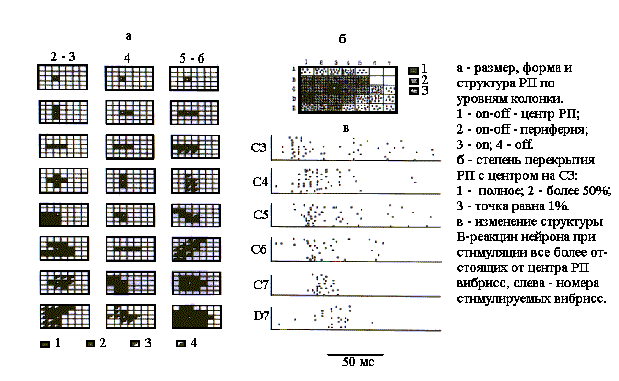
Рис. 2. РП нейронов разных уровней бочонковой колонки С1.
Нейроны поверхностных (2-3) и глубоких (5-6) слоев имели многовибриссные РП, которые всегда были сплошными. У 23% нейронов РП имели вытянутую, чаще всего вдоль, но иногда и поперек вибриссковых рядов, форму. В последнем случае угол наклона РП к рядам вибрисс варьировал у разных нейронов, но чаще всего был прямым. Однородную структуру РП с on-off-ответами имели 31%, а с on-ответами - 8% нейронов. Большинство нейронов имели неоднородные РП: с on-off-ответами в центре и с оn-ответами по краю - 23%, а с on-off-центром и off-краем - 38% нейронов. При этом у 70% нейронов центр РП располагался эксцентрически, т.е. находился вблизи края В-зоны РП. Средние значения амплитудных порогов у нейронов с многовибриссными РП были достоверно (Р<0,01) выше, чем у одновибриссных: на уровне поверхностных слоев - 9,4+0,3 мкм, а в глубоких слоях - 13,0+0,8 мкм. Нейроны, РП которых перекрывали почти все контралатеральное поле вибрисс, имели амплитудные пороги около 30 мкм.
РП нейронов, принадлежащих одной и той же бочонковой колонке, существенно перекрывались. Центры РП данной выборки нейронов всегда совпадали, поскольку это было исходной предпосылкой для отнесения регистрируемых нейронов к бочонковой колонке С3. Наложением картированных РП произведена количественная оценка степени их перекрытия во всей выборке нейронов, представляющих колонку С3 (рис.2,б). Анализ показал, что все вибриссы контралатеральной стороны в той или иной мере адресованы нейронам данной колонки. При этом степень перекрытия РП градуально уменьшилась к краям вибрисскового поля до 5%. Однако, подзона с более чем 50-процентным перекрытием РП, заштрихованная на рис.2б, имела симметричную и вытянутую вдоль ряда С форму. Таким образом, суммарное РП нейрональной колонки С3 ориентировано вдоль одноименного ряда вибрисс, в плоскости которого совершаются их ощупывающие движения, и имеет значительное перекрытие с РП нейронов соседних бочонковых колонок этого, а также верхнего и нижнего рядов.
На рис.2,в в качестве примера более общей закономерности показано удлинение ЛП, уменьшение частоты первичного разряда, а также перестройка структуры В-реакции нейрона, зарегистрированного на уровне слоя 4, при поочередной стимуляции вибрисс, отстоящих на все большем расстоянии от центра РП. В среднем по выборке (N=52) ЛП при смещении стимула от центра на периферию уменьшался с 9,0+0,3 мс до 19,1+1,2 мс. Уменьшалась при этом также и частота первичного разряда в первые 50 мс после отметки стимула с 132+21 имп/с до 41+8 имп/с. Все это в совокупности свидетельствует об уменьшении функциональной эффективности синаптических влияний на колонку по направлению к периферии РП ее нейронов.
На рис.1,б показана структура В-зоны РП нейрона, зарегистрированного на глубине 1100 мкм и принадлежащего слою 5, при разных амплитудах тактильного стимула, а также соответствующие им точечные ПСГ (отклонялась вибрисса С3). При 10-пороговой амплитуде (90 мкм) В-зона РП данного нейрона состояла из 14 вибрисс, 6 из которых вызывали on-off-ответы и находились в центре РП, а 8 периферических вибрисс - on-ответы. При уменьшении амплитуды количество вибрисс в В-зоне РП уменьшалось вначале незначительно и, в основном, за счет периферии. Значительное уменьшение площади РП наблюдалось у этого и других нейронов при амплитудах 30 и 22 мкм и совпадало с исчезновением в структуре В-реакции нейрона на отклонение даже центральной вибриссы компонентов вторичной активации.
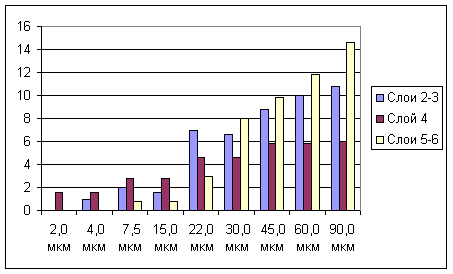
Рис. 3. Зависимость размеров В-зоны РП нейронов разных уровней бочонковой колонки С1 от амплитуды сгибания вибриссы.
На рис.3 показан характер изменения средних размеров В-зоны РП нейронов трех уровней бочонковой колонки С3 от амплитуды сгибания соответствующей вибриссы. Следует отметить, что размер РП у нейронов слоя 4 существенно уменьшался только при амплитуде 22 мкм, тогда как у нейронов других уровней - уже при более высоких амплитудах (45 и 30 мкм), что коррелировало с их более высокими амплитудными порогами. В диапазоне околопороговых амплитуд РП нейронов слоя 4 становилось равными по размеру с РП нейронов других слоев.
Размер и структура РП обнаружили также отчетливую зависимость от уровня возбуждения бочонковой колонки. В модельной серии опытов общий уровень возбудимости мозга крыс повышался посредством внутрибрюшинного введения кофеина в дозе 100 мг/кг /24/. Так, на рис.4,А представлены результаты картирования РП нейрона, зарегистрированного в опыте N 13 на глубине 1200 мкм в бочонковой колонке С3. ПСГ в части А рисунка, представляющие центральную часть РП, построены по 32 реализациям и показывают наибольшую эффективность специфического входа в данную колонку от вибриссы С3.
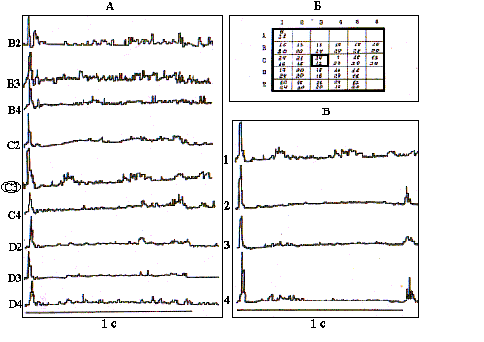
Рис. 4. Перестройка структуры В-зоны РП нейрона при повышении общего уровня возбудимости коры после введения кофеина.
А - ПСГ реакций нейрона на сгибание соответствующих вибрисс, С3 - центр РП; Б - карта РП, цифры вверху - накопление импульсов в первичную фазу, внизу - ЛП (мс) максимума ПСГ; В - формирование off- реакции на стимуляцию центра РП после введения кофеина: 1- контроль, 2- через 10 мин, 3- 20 мин, 4- 40 мин.
По всему РП в контроле данный нейрон реагирует возбуждением только на включение стимула. В части Б данного рисунка изображена полная карта РП этого нейрона со значениями максимумов в фазе первичного возбуждения и их ЛП, которые дают представление о более тонкой структуре РП. С учетом представленных показателей в РП данного нейрона также можно выделить горизонтальную в пределах рядов С и D ориентационную избирательность, проявляющуюся, прежде всего, в более высокочастотных первичных разрядах и, в меньшей мере, в их ЛП.
Через 10 мин после введения кофеина нейрон начинает реагировать также и на выключение стимула, приложенного в центре РП (рис.4,В). Со временем интенсивность первичного возбуждения на включение и выключение стимула уменьшается (к 20-й минуте), а затем к 40-й минуте вновь возрастает. Кроме того, постепенно восстанавливается вторичная активация, подавляемая на начальных этапах действия кофеина. Данный пример является весьма типичным для всей выборки исследованных нейронов (N=20). Нейроны бочонковых колонок, представляющих в С1 коры мозга крыс латеральные ряды вибрисс, имели во всех исследованных случаях односторонние РП, располагающиеся с существенным перекрытием по всему вибриссковому полю контралатеральной стороны мордочки крысы. Нейроны в представительстве самого медиального ряда вибрисс А, напротив, всегда имели билатеральные РП (N=20). В бочонковых колонках ряда В с равной вероятностью встречались и односторонние и двусторонние РП (N=10). Размеры билатеральных РП соответствовали односторонним. Их структура в общих чертах характеризовалась наличием более детерминированной по ответоспособности центральной части и периферии с низкой проявляемостью реакций на серию повторяющихся стимулов. Ипсилатеральная часть РП соответствовала периферии, однако, в ней также выделялась центральная вибрисса, симметричная центральной в контралатеральной части РП, которая давала более интенсивные и с большей вероятностью реализующиеся импульсные реакции. Кроме того, ипсилатеральная часть РП всегда ограничивалась лишь двумя медиальными рядами вибрисс - А и В. Структура импульсных реакций нейронов при стимуляции ипсилатеральных вибрисс также была фазной, с четкой тормозной паузой между первичным и вторичным возбуждением, свидетельствующей об участии системы ассоциативных связей в обработке ипсилатеральной специфической афферентации.
Контралатеральная часть РП нейронов в представительстве медиальных вибрисс, как правило, не включала в свой состав самые нижние вибриссы рядов D и Е. На рис. 5 представлено РП нейрона слоя 5 бочонковой колонки А2. Оно включает в свой состав 9 вибрисс в медиальных рядах А и В контралатеральной стороны и две вибриссы (А2 и В2) на ипсилатеральной стороне мордочки крысы. В табличках представлены значения максимумов ПСГ, построенных по 32 реализациям, а также их ЛП и длительность тормозной паузы (последние два показателя в мс). Видно, что реакции на отклонение центральных вибрисс в левом и правом полуполях РП характеризуются наибольшей интенсивностью первичного и структурированностью вторичного возбуждения. Показательны также значения тормозной паузы, максимальные для центральной вибриссы, а также значения ЛП реакций, которые в пределах всего ряда А не различались, но были достоверно (P<0,01) короче, чем у В-реакций в ответ на стимуляцию вибрисс ряда В одноименной стороны. Следует также отметить низкую проявляемость нейронных реакций в ответ на повторную стимуляцию вибрисс ипсилатеральной части РП, которая не превышала по выборке 60%, что предполагает кооперативный характер деятельности нейронов с билатеральными РП при восприятии тактильных стимулов.
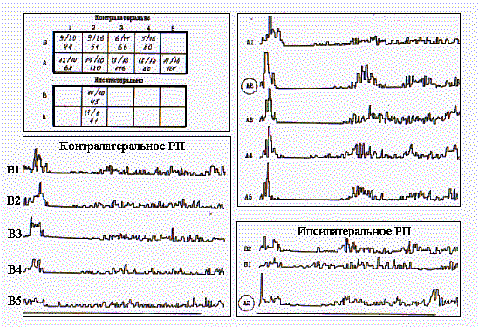
Рис.5. Билатеральное РП нейрона бочонковой колонки в представительстве медиального ряда вибрисс (объяснения в тексте).
Параметры нейронных реакций на контралатеральную и ипсилатеральную стимуляцию.
В данной серии опытов исследованы реакции нейронов бочонковых колонок представительства латеральных вибрисс в С1 коры мозга крыс на контралатеральную и ипсилатеральную стимуляцию вибрисс, а также находящейся под ними шерсти. В последнем случае в качестве топографических ориентиров при картировании РП использовались вибриссы. На рис.6,а представлены графики послойного распределения 91 нейрона, зарегистрированных в данной серии, которые реагировали первичным возбуждением на отклонение центральных в РП вибрисс контралатеральной стороны мордочки крысы (КВ-нейроны) и 54 из них, реагирующих, кроме того, на тактильную стимуляцию шерсти ипсилатеральной стороны. Мы назвали такие нейроны билатеральными (БВ-нейроны). Следует при этом отметить, что КВ-нейроны при стимуляции шерсти ипси-стороны давали такие же импульсные реакции, только более высокочастотные и коротколатентные, как и при стимуляции вибрисс контралатеральной стороны. Поэтому этот тип реагирования нейронов мы не стали выделять в качестве самостоятельного, тем более что при стимуляции шерсти не удается избежать одновременной стим