


Лигнин
Введение.
В технологии переработки древесной зелени под хвойной древесной зеленью понимают – хвойную лапку, то есть охвоенные побеги, таким образом, хвойная древесная зелень представляет собой смесь хвой, коры и древесины. Отработанная древесная зелень представляет собой остаток после экстракции древесной зелени бензином и обработки острым паром. Фитосорб представляет собой остаток после экстракции древесной зелени бензином, обработки острым паром, экстракции разбавленной щелочью и отмывки щелочи водой.
Теоретическая часть.
Раздел 1.
1.1. Древесная зелень.
Листья древесных растений выполняют ряд важных функций, что находит отражение в их строении и химическом составе. Они осуществляют фотосинтез, испарение воды (транспирацию) и газообмен (поглощение и выделение диоксида углерода и кислорода).
Поглощаемый из атмосферы СО в ассимиляционных тканях листа превращается в органические соединения, которые затем передаются в остальные органы дерева. В этих процессах синтеза листья используют солнечную энергию, т.е. происходит фотосинтез. Побочным продуктом фотосинтеза является кислород, выделяемый в атмосферу. Дополнительную энергию и целый ряд химических соединений древесные растения получают в результате дыхания, в ходе которого происходит окисление органических веществ. В противоположность фотосинтезу при дыхании поглощается кислород, а выделяется СО как один из конечных продуктов окисления.
Листья очень разнообразны по форме и строению, но их можно подразделить на две группы: листья покрытосеменных растений, в том числе лиственных деревьев и других растений этого отдела, используемых в качестве сырья для химической переработки; листья голосеменных растений (хвойных деревьев). Основной частью листа, в которой осуществляются его функции, является листовая пластинка. У хвойных деревьев форма листовой пластинки чаще всего игловидная или чешуевидная, в поперечном сечении плоская, трех- или четырехгранная, полукруглая, овальная. Длина хвоинок у ели обыкновенной 1,5...2,0 см, у кедра сибирского 8...13 см, а у некоторых пород, например, у сосны длиннохвойной (Pinus palustris) может достигать 20...30 см (у молодых растений даже до 45 см). Игловидные листья обычно располагаются на укороченном побеге пучком (мутовкой). Число хвоинок на побеге составляет 1...5. Пластинчатые листья у лиственных деревьев значительно более разнообразны по форме и размерам. Продолжительность жизни листьев у листопадных деревьев и однолетних растений несколько месяцев, а у вечнозеленых - до 1,5...5 и даже 15 лет (у ели Шренка до 28 лет), после чего они заменяются новыми.
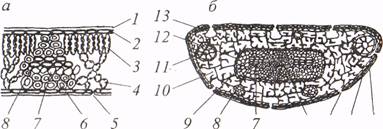
Листья имеют покровные, основные и проводящие ткани. Снаружи листья покрыты первичной покровной тканью - эпидермой. Основные ткани, т.е. ткани, составляющие основную массу листа, называют мезофиллом. Мезофилл состоит из паренхимных клеток с хорошо развитой пигментосодержащей системой пластид и служит ассимиляционной тканью. Проводящие ткани листьев - ксилема и флоэма - образуют проводящие пучки. Структура игольчатого листа имеет свои особенности. На рис. 1.1 в качестве примера изображены поперечные срезы листа березы и хвои сосны.
Эпидерма состоит из одного слоя (реже нескольких) сплюснутых таблетчатых клеток, плотно сцепленных между собой. Снаружи эпидерма покрыта гидрофобной кутикулой, которая обычно состоит из трех слоев. Внешний слой образован воском, представляющим смесь соединений с длинноцепочечными молекулами (сложных эфиров высших жирных кислот и высших алифатических спиртов, свободных высших жирных кислот, гидроксикислот, алифатических углеводородов, спиртов и кетонов). Его называют эпикутикулярным воском. Под ним лежит слой заключенного в воск кутина. Кутин является основным полимером кутикулы. Он сходен по строению с суберином, т.е. это сложный полиэфир сетчатого строения, образованный преимущественно и кислотами (насыщенными и одноненасыщенными жирными кислотами, насыщенными и одноненасыщенными гидроксикислотами, которые могут дополнительно иметь одну или две гидроксильные группы). Внутренний слой кутикулы кроме кутина и воска содержит полисахариды.
У игольчатых листьев под эпидермой располагается плотный слой клеток гиподермы, подобных волокнам и имеющих толстые стенки. Гиподерма относится к склеренхимным (механическим) тканям (гиподермальная склеренхима). У различных хвойных эта ткань варьируется по содержанию. В эпидерме имеются многочисленные устьица (десятки и даже сотни на 1 мм2), через которые происходят газообмен и транспирация. Под устьицами у хвои слой гиподермы также прерывается. Эпидерма (вместе с гиподермой) служит механической опорой для мезофилла и других тканей листа.
Мезофилл в пластинчатых листьях подразделяется на палисадную (столбчатую) паренхиму с вытянутыми клетками и губчатую паренхиму с крупными межклетниками. В хвое клетки мезофилла обычно плотно сжаты с образованием складок в клеточных стенках. Такой тип фотосинтезирующих клеток называется складчатой паренхимой. Паренхимные клетки мезофилла содержат пластиды, называемые хлоропластами - тельца дисковидной формы (размером 3...10 мкм) зеленого цвета, расположенные по периферии цитозоля вплотную к плазмалемме. Хлоропласта способны синтезировать зеленые пигменты - хлорофиллы и желтые - каротиноиды. Подобный тип паренхимы называют хлоренхимой. Межклетники мезофилла обеспечивают интенсивный газообмен между наружным воздухом и фотосинтезирующей тканью. Кроме того, с поверхности клеток вода сначала испаряется в межклетники, а затем через устьица транспирируется. При ослаблении напора воды в клетках, образующих устьица, они схлопываются. Эпидерма вместе с кутикулой защищает мезофилл от значительной потери воды.
В мезофилле хвои проходят смоляные ходы, выстланные тонкостенными эпителиальными клетками и окруженные снаружи склеренхимными волокнами с утолщенными одревесневшими стенками. Число ходов зависит от древесной породы и от условий произрастания. В хвое сосны смоляные ходы особенно развиты, тогда как у тиса вообще отсутствуют (так же как и гиподерма). Со смоляными ходами побегов смоляные ходы хвои контактируют через систему паренхимных клеток.
Лист пронизан жилками, которые включают один или несколько проводящих пучков, образованных клетками первичной ксилемы и первичной флоэмы. Вдоль жилок группируются склеренхимные волокна. Флоэмная паренхима развита сильнее ксилемной. Ксилема обращена к морфологически верхней стороне листа, флоэма - к нижней. Ксилема образована трахеидами. В паренхимных клетках ксилемы накапливается крахмал. Флоэма состоит из паренхимных клеток и ситовидных трубок. Во флоэме имеются альбуминовые клетки с густой цитоплазмой, богатой белками. Проводящие ткани хвои объединены в проводящий цилиндр, который обычно содержит два проводящих пучка и отграничен от клеток мезофилла эндодермой. Проводящие пучки окружены трансфузионной тканью, состоящей из живых паренхимных клеток с неодревесневшими стенками и трахеид с тонкими, но одревесневшими стенками. Трансфузионные трахеиды и клетки трансфузионной паренхимы образуют две взаимопроникающие системы. Проводящие пучки иногда отделяются от трансфузионной ткани прерывистым слоем склеренхимы. Трансфузионная ткань осуществляет передвижение органических веществ и воды между проводящим пучком и мезофиллом. Эндодерма, окружающая трансфузионную ткань, состоит из относительно толстостенных клеток, иногда содержащих крахмал. Данный слой клеток четко выражен в хвое деревьев сем, сосновых и некоторых других хвойных пород. Вторичные стенки клеток эндодермы пропитаны суберином и лигнифицированы.
Функции хвои (фотосинтетическая активность), смолистость и механические свойства зависят от количественного соотношения ассимиляционных, проводящих и механических тканей, которое, в свою очередь, зависит от древесной породы, по колеблется даже в пределах одного и того же рода. Так, проводящая система наиболее развита в хвое различных видов сосны и слабо у ели. Лиственница и пихта занимают промежуточное положение.
Развитие листа, в том числе и хвои - сложный процесс, связанный с делением меристематических клеток, дальнейшим ростом вновь образовавшихся клеток, их дифференциацией и развитием проводящей системы. Через определенное время происходит опадение листьев перед наступлением зимы. У хвойных деревьев одновременного ежегодного опадения листьев нет (за исключением лиственницы), но периодически часть хвои отмирает и заменяется новой.
Листья деревьев, главным образом хвою, используют в лесохимии как сырье, заготавливаемое в виде так называемой древесной зелени. В технологии переработки древесной зелени под хвойной древесной зеленью понимают хвойную лапку, т.е. охвоенные побеги. Для характеристики такого сырья его разделяют на хвою, кору и древесину. Наиболее ценные части, богатые биологически активными веществами, - хвоя и кора.
Анатомическое строение древесины, входящей в древесную зелень, в основных деталях не отличается от строения древесины ствола (стебля). По определению Яценко-Хмелевского, побег представляет собой облиственный (охвоенный) стебель. От ствола дерева отходят ветви первого порядка, от них ветви второго порядка и т.д. вплоть до побегов. Верхушки побегов содержат меристематическую ткань (верхушечную, или апикальную, меристему). В этих верхушках и возникают листья. В побегах (без хвои) очень велика доля паренхимных клеток за счет значительного объема (до 70%) молодой коры, что и приводит к высокому содержанию биологически активных веществ.
Практическое использование хвойной древесной зелени обусловлено ее особым химическим составом, наличием не только биологически активных соединений (витамина С, каротина, ферментов, гормонов и др.), но и ценных питательных веществ (протеина в количестве 8... 15%, жиров- 5...8%, легкоперевариваемых углеводов-до 3%). Это позволило разработать технологию промышленной заготовки и переработки древесной зелени с получением хвойной витаминной муки и других кормовых добавок, пихтового эфирного масла, хвойного экстракта, хлорофилло-каротиновой пасты, других биологически активных препаратов. Перспективной считают переработку и лиственной древесной зелени.
В химическом составе хвойной древесной зелени можно выделить те же самые основные высокомолекулярные компоненты древесины - целлюлозу, гемицеллюлозы, лигнин. Однако их доля существенно ниже, чем в древесине той же древесной породы, и значительно выше доля экстрактивных веществ, состав которых более разнообразен
6521
Рис. 1.1. Поперечные срезы листа березы (а) и сосны (б): 1 - эпидерма; 2 - кутикула; 3 - палисадная (столбчатая) паренхима; 4 - губчатая паренхима; 5 - устьице; 6 -склеренхимныс волокна; 7 - флоэма; 8 - ксилема; 9 - трансфузионная ткань; 10 - складчатая паренхима; 11 - смоляной ход; 12 - эндодерма; 13 - гиподерма
1.2 Экстрактивные вещества древесной зелени.
Основные компоненты экстрактивных веществ древесины присутствуют и в древесной зелени. Однако для зелени характерно наличие некоторых специфических веществ, таких как фотосинтетические пигменты, витамины и сложные липиды.
Эфирные масла хвои отличаются чрезвычайным разнообразием присутствующих терпенов и терпеноидов. Выход эфирных масел из хвои обычно невелик (0,2...3,0%), у хвои пихт - до 5%. В их составе превалируют монотерпены, массовая доля которых достигает 60...90%. В монотерпеновой фракции хвои различных видов сосны, произрастающих в России, главными компонентами являются -пинен и карен-3. Состав эфирного масла из древесной зелени зависит от соотношения хвои и побегов. В России в значительных количествах из древесной зелени пихты сибирской получают пихтовое масло. В его состав входит около 100 компонентов, главным образом, монотерпенов и их производных.
Среди веществ, растворимых в воде, преобладают таннины и углеводы. Хвоя содержит довольно много таннинов: еловая до 10%, а сосновая около 3...4%. Содержание моно- и олигосахаридов подвергается сезонным изменениям, увеличиваясь зимой и снижаясь летом. Среди олигосахаридов сосны обыкновенной преобладают сахароза и рафиноза, выделены также стахиоза и мелибиоза (схема 1.2). В водных экстрактах древесной зелени содержатся органические кислоты, флавоноиды и другие фенольные соединения и их гликозиды, водорастворимые белки и группа водорастворимых витаминов. Количество веществ, экстрагируемых из хвои органическими растворителями, и их состав определяются природой растворителя. Неполярный растворитель - петролейный эфир - извлекает из еловой, пихтовой и сосновой хвои до 8... 14% ее массы, малополярный диэтиловый эфир - 10.. .22%, а растворители, способные к образованию водородных связей (этанол, изопропанол), - до 30...40%. Состав этой группы веществ весьма разнообразен: пигменты, витамины, липиды (простые липиды, глико- и фосфолипиды).
1.3 Фотосинтетические пигменты.
Древесная зелень отличается от остальной биомассы дерева наличием большого числа клеток, содержащих хлоропласты, главным образом, клеток палисадной и губчатой тканей мезофилла листа и в меньших количествах других зеленых тканей. Обычно хлоропласты окрашены в зеленый цвет, и именно их присутствию зелень обязана своим названием. Они содержат фотосинтетические пигменты двух классов - хлорофиллы и каротиноиды, поглощающие свет разных длин волн. Хлорофиллы относятся к зеленым пигментам и поглощают свет в синей и красной областях видимой части спектра при длинах волн соответственно около 450 нм и 650...700 нм. Каротиноиды - желтые и оранжевые пигменты, поглощающие свет в области 400...500 нм.
Пигменты входят в состав пигментных систем в виде хромопротеинов, т.е. пигмент- белковых комплексов. Компоненты в этих комплексах соединены только межмолекулярными связями. Поэтому пигменты можно экстрагировать из измельченной древесной зелени органическими растворителями. Это могут быть и неполярные растворители, но более полное извлечение достигается с помощью ацетона или этанола.
Во всех фотосинтезирующих растениях обнаружен хлорофилл а, содержание которого превышает содержание других пигментов. Он является самым важным пигментом, так как образует реакционные центры, участвующие в световой фазе фотосинтеза. Другие формы хлорофиллов, а также каротиноиды рассматриваются как вспомогательные, или сопутствующие, пигменты. Функция каротиноидов не ограничивается ролью светособирающих пигментов. Они также защищают ткани от окисления кислородом на свету.
Хлорофиллы. Зеленые пигменты растений - хлорофиллы имеют определенное родство с гемом (гемином) - красным пигментом крови. И гем и хлорофиллы относятся к порфиринам. Порфирины - важнейшие органические компоненты биологических систем, имеющие в качестве основной структурной единицы гетероцикл пиррола. Порфирины содержат в молекуле макроцикл порфина - циклическую тетрапиррольную структуру с метиленовыми мостиками. Порфирины различаются боковыми заместителями и способны образовывать комплексы (хелатные соединения) с металлами. Хлорофилл - это Мg-порфириновый комплекс, а гем - Fе-порфириновый. Биологическая активность порфиринов зависит как от металла, образующего комплекс, так и от набора и расположения заместителей - метальных, этильных, винильных групп и, главным образом, остатков пропионовой кислоты. В основе молекул хлорофиллов лежит гетероциклическая структура форбина содержащая по сравнению с порфином дополнительное кольцо циклопентана, образующееся при циклизации с участием одного из остатков пропионовой кислоты. Второй остаток связан в виде сложного эфира с ациклическим дитерпеновым спиртом фитолом С Н ОН. Одно из ниррольных колец (в положениях 7,8) гидрировано. В растениях, в том числе древесных, кроме хлорофилла а присутствует хлорофилл Ь с формильной группой в положении 3 вместо метильной.
В молекуле хлорофилла тетрапиррольный макроцикл представляет собой систему сопряженных двойных связей с делокализованной электронной плотностью, т.е. в этой системе не фиксировано положение двойных связей. Из-за этого не фиксировано и положение двух ковалентных и двух координационных (донорно-акцепторных) связей центрального атома магния с четырьмя атомами азота. На схеме 1.3 приведены две из возможных резонансных структур хлорофилла. Заряд на атоме магния (2+) компенсируется двумя избыточными электронами, распределенными между четырьмя атомами азота, и молекула хлорофилла электрически нейтральна.
Хлорофиллы, как и другие порфирины, образуются сложным многостадийным процессом биосинтеза из аминокислот с участием специфических ферментов.
Массовая доля хлорофиллов в хвое может варьироваться в зависимости от возраста и условий произрастания деревьев от 0,04 до 1,2%, но обычно редко превышает 0,3%. В древесной зелени содержание хлорофилла будет определяться долей в ней хвои; так, в хвое пихты сибирской массовая доля хлорофилла составляет 0,9%, а в побегах (без хвои) - 0,2%.
Хлорофиллы неустойчивы к действию кислорода (особенно на свету), разбавленных растворов кислот и щелочей. При действии фермента хлорофиллазы отщепляется фитол. Аналогичным образом действуют щелочи. При действии кислот магний замещается на водород, а при более длительном воздействии кислот, кроме того, гидролизуется сложноэфирная связь с освобождением фитола. Эти свойства хлорофиллов имеют значение для химической переработки древесной зелени.
Каротиноиды. Желто-оранжевые каротиноидные пигменты рассматривают как производные тетратерпенов. Окраска каротиноидов обусловлена наличием в молекуле длинного ряда сопряженных двойных связей. Каротиноиды, представляющие собой углеводороды, называют каротинами, а их кислородсодержащие производные - ксантофиллами. Каротиноиды липофильны - растворимы в жирах (маслах), в диэтиловом эфире. Растворимость в других растворителях определяется строением каротиноида.
Известно большое число каротиноидов. В зеленых листьях содержится главным образом р-каротин (схема 1.4). Каротины являются соединениями, из которых в организмах человека и животных образуется витамин А, т.е. их рассматривают как провитамин А. Витамин А представляет группу природных соединений - производных (3-ионона, важнейшими представителями которых служат ретинол, ретиналь и ретиноевая кислота (см. схему 1.4). В животных организмах под действием ферментов молекула - карогина превращается в две молекулы ретиналя, которые затем восстанавливаются до ретинола и незначительно окисляются до ретиноевой кислоты. Массовая доля каротиноидов в зеленых листьях составляет 0,07. . .0,2%.
1.4. Витамины.
К витаминам относят низкомолекулярные органические соединения различной химической природы, необходимые для осуществления биохимических и физиологических процессов. Организмы человека и животных практически не способны синтезировать витамины и должны получать их с растительной пищей. Витамины подразделяют на водорастворимые и жирорастворимые. В древесной зелени присутствуют представители различных групп витаминов.
Из жирорастворимых витаминов древесная зелень содержит витамины групп Е (токоферолы), О (кальциферолы) и К (производные нафтохиноиа). В зелени присутствуют почти все водорастворимые витамины: витамин С (аскорбиновая кислота), по содержанию которого хвою сосны и ели можно приравнять к ягодам черной смородины; витамины группы В - тиамин (В1), рибофлавин (В2), пиридоксин (Вб), пантотеновая кислота (Вз); фолацин (фолиевая кислота и ее производные, т.е. витамин В9); ниацин (никотиновая кислота и ее производные, т.е. витамин РРЦ).
Древесная зелень богата биологически активными веществами. Кроме собственно витаминов она содержит большое количество уже упоминавшегося провитамина А, ряд стеринов - провитаминов D. Также в зелени содержатся витаминоподобные вещества (бифлавоноиды - витамин Р, циклические спирты инозиты и др.), которые по своим функциям в животных организмах близки или к витаминам, или к другим незменимым пищевым веществам (незаменимым жирным кислотам и аминокислотам).
Древесная зелень содержит, главным образом в связанном виде, все незаменимые кислоты, а также незаменимые полиненасыщенные кислоты — линолевую и линоленовую.
1.5. Липиды.
Термин липид в определенной мере условен, поскольку под липидами понимают жироподобные вещества, входящие в состав всех живых клеток. Иногда к липидам относят различные по строению органические соединения, присутствующие в живых тканях, не растворимые в воде и извлекаемые из тканей неполярными органическими растворителями (диэтиловый эфир, бензол, хлороформ). Однако при таком подходе в состав липидов наряду с жирами попадают самые разные по своей природе соединения: терпены и терпеноиды, смоляные кислоты, каротиноиды, хлорофиллы, витамины и др. Поэтому часто при отнесении соединений к липидам учитывают и химическое строение. В соответствии с химическим строением выделяют три группы собственно липидов: жирные кислоты и продукты их ферментативного окисления (проста-гландины и другие гидроксикислоты); глицеролипиды (содержат в молекуле остаток глицерина); липиды разного состава, не содержащие остатка глицерина и не относящиеся к липидам первой группы (некоторые фосфолипиды и гликолипиды, диольные липиды, стерины и воски). Существуют и другие системы классификации липидов. Липиды создают в растительной ткани энергетический резерв, образуют защитные покровные ткани, служат запасными питательными веществами, входят в состав клеточных мембран. Липиды также подразделяют на простые и сложные. Простые (нейтральные) липиды хорошо растворяются в неполярных углеводородных органических растворителях (петролейный эфир, бензол), а сложные (полярные) - в полярных растворителях (этанол). При хроматографическом разделении нейтральные (неполярные) липиды можно элюировать неполярными растворителями, тогда как для элюирования полярных липидов требуются высокополярньге растворители или смеси растворителей. К простым липидам относят жиры (триацилглицерины) и воски, к сложным - гликолипиды и фосфолипиды. Гликолипиды представляют собой гликозиды, у которых в роли агликона выступают преимущественно диацилглицерины. Для хлоропластов характерно наличие гликолипидов, представляющих собой главным образом моно- и дигалактозилдиацилглицерины с жирнокислотными остатками, отличающимися высокой ненасыщенностью (схема 1.5, а).Фосфолипиды -это диэфиры ортофосфорной кислоты. В фотосинтезирующих тканях растений найдены различные глицерофосфолипиды, которые можно рассматривать как эфиры фосфатидовых кислот (фосфатиды), этерифицированные холином, глицерином, этаноламином, инозитом (см. схему 1.5, б). Глицерофосфолипиды могут образовывать комплексы с белками (липопротеиды). Простых липидов в древесной зелени содержится больше, чем гликолипидов, которых в свою очередь почти в два раза больше, чем фосфолипидов. В простых липидах хвои сосны присутствуют главным образом диеновые кислоты (линолевая и др.) и моноеновые, которых в два раза больше триеновых и насыщенных кислот и значительно больше, чем тетраеновых. Гликолипиды отличаются повышенным содержанием триеновых кислот. В фосфолипидах превалируют диеновые и насыщенные кислоты. Низкомолекулярных С … С -кислот в гликолипидах и фосфолипидах почти нет, в отличие от простых липидов.
Раздел 2.
2.1. Общие понятия о лигнине.
Лигнин - это ароматическая часть древесинного вещества. С аналитической точки зрения лигнин рассматривают как ту часть древесины, которая получается в виде нерастворимого «негидролизуемого» остатка после удаления экстрактивных веществ и полного гидролиза полисахаридов концентрированной сильной минеральной кислотой. В действительности лигнин нельзя считать негидролизуемым полимером, поскольку при действии кислот в лигнине могут гидролизоваться углерод-кислородные связи, но сохраняются существующие в нем углерод-углеродные связи и образуются новые.
Массовая доля лигнина в древесине хвойных пород составляет в среднем 27...30%, а в древесине лиственных пород - 18. ..24%. В отличие от целлюлозы и других полисахаридов выделенный из древесины лигнин не является индивидуальным веществом, а представляет собой смесь ароматических полимеров родственного строения.
2.2 Структурные единицы лигнина.
Лигнин как полимер состоит из фенилпропановых структурных единиц (мономерных составляющих звеньев) - ФПЕ, обозначаемых сокращенно С6-С3, или единицы С9. Лигнин хвойных пород, появившихся в эволюции раньше лиственных, имеет структурных единиц (I). В состав лигнина лиственных пород, кроме гваяцилпропановых единиц, в значительных количествах входят сирингилпропановые единицы (II). Гваяциллропановые единицы (G-единицы) рассматривают как производные пирокатехина, а сирингилпропановые единицы (S-единицы) - как производные пирогаллола (схема 1.6). В соответствии с основным типом структурных единиц лигнины хвойных пород относят к гваяцильным лигнинам, а лигнины лиственных пород - к гваяцилсиригильным лигнинам. Кроме гваяцилпропановых единиц в состав хвойных лигнинов входят в небольшом количестве, а в состав лиственных лигнинов в еще меньшем n-гидроксифенилпропановые единицы (III). По сравнению с лигнинами древесных пород гидроксифенилпропановые единицы (Н-единицы) значительно большую долю составляют в лигнинах однодольных растений (трав, злаков).При нитробензольном окислении фенилпропановые единицы дают ароматические альдегиды. Гваяцилпропановые единицы окисляются в ванилин, сирингилпропановые - в сиреневый альдегид, гидроксифенилпропановые единицы в n-гидроксибензальдегид. По соотношению альдегидов судят о соотношении в лигнинах структурных единиц разного типа. Так, у лигнина ели (Рicea abies) соотношение единиц G:S:Н = 94:1:5, а у лигнина сосны (Pinus taeda) 86:2:12. Все лигнины, даже в пределах одной клеточной стенки, гетерогенны. Поэтому приводимые в литературе данные по соотношению ФПЕ разного типа следует рассматривать только как средние. Лиственные лигнины более гетерогенны, чем хвойные, и доля сирингильных единиц может составлять от 20 до 60%, а иногда и выше. Например, в лигнине бука (Fagus sylvatica) соотношение единиц G:S:Н по разным данным составляет 56:40:4; 49:46:5. У травянистых растений доли сирингильных и гидроксифенильных единиц колеблются в широких пределах.
В химии лигнина принята своя нумерация атомов углерода бензольных колец в ФПЕ, отличающаяся от нумерации принятой в органической химии для фенолов, как показано на примере гваяцилпропановой единицы (см. схему 1.6). Атомы пропановой цепи обозначают греческими буквами. При написании формул структурных единиц лигнина в схемах реакций иногда пользуются сокращенными изображениями пропановых цепей:
2.3 Природный лигнин и препараты лигнинов.
Различают природный лигнин (протолигнин) - лигнин, находящийся в древесине, и препараты лигнина - лигнины, выделенные из древесины. Природный лигнин в древесине неоднороден. Основная его масса имеет трехмерную сетчатую структуру и вследствие этого не растворяется в растворителях. Для перевода природного лигнина в растворимое состояние с целью удаления (делигнификации) или выделения из древесины (получения растворимого препарата) требуется разрушение сетки с помощью довольно жесткого химического воздействия («химическое растворение»). Как у всех сетчатых полимеров, понятие макромолекулы для природного лигнина теряет смысл.
Среди выделенных лигнинов различают препараты нерастворимых и растворимых лигнинов. Нерастворимые лигнины сохраняют сетчатую структуру. Растворимые лигнины, представляющие собой фрагменты сетки, имеют разветвленную структуру с высокой степенью разветвленности и неоднородны по молекулярной массе.
Любой лигнин, как лиственных, так и хвойных древесных пород, представляет собой гетерополимер. Кроме различий в типе составляющих фенилпропановых единиц (G, S или Н), в пропановых цепях единиц одного и того же типа в пределах макромолекулы или сетки содержатся разные функциональные группы (гидроксильные, карбонильные, карбоксильные, двойные связи) и присутствуют связи разного типа с другими единицами. Следовательно, лигнин имеет высокую степень химической неоднородности. Разветвленные макромолекулы растворимых лигнинов и протяженные участки цепей в сетчатой структуре, в отличие от таких важнейших биополимеров растительных и животных тканей, как целлюлоза и белки, имеют нерегулярное строение.
Лигнин - это органический гетероцепной кислородсодержащий полимер, но в отличие от полисахаридов, относящихся к полиацеталями, у лигнина отсутствует единый тип связи между мономерными звеньями. Наряду с углерод—кислородными (простыми эфирными) связями С-О—С присутствуют и углерод-углеродные связи С—С между звеньями, характерные для карбоцепных полимеров. В структурных единицах лигнина содержатся различные полярные группы и в том числе способные к ионизации (кислые) фенольные гидроксилы и в небольшом числе карбоксильные группы, вследствие чего лигнин является полярным полимером, проявляющим свойства полиэлектролита. Лигнин - аморфный полимер, как природный, так и выделенный. Из-за высокой степени разветвленности макромолекулы выделенных растворимых лигнинов имеют глобулярную форму и такие препараты представляют собой порошки. В лигнине, благодаря наличию большого числа гидроксильных и других полярных групп, значительно развиты водородные связи (внутри- и межмолекулярные).
Лигнин очень рано образуется в растительных клетках. В первые дни после отделения новой клетки ксилемы от камбиальной клеточная стенка еще не содержит лигнина. Через сравнительно небольшой период времени (у некоторых растений даже на второй-третий день) клеточные стенки начинают давать качественные (цветные) реакции на лигнин (см. 12.3). Процесс лигнификации имеет большое значение для жизни дерева и сыграл важную роль в эволюции растений. Именно лигнификация растительных тканей позволила осуществить переход растений от водного к наземному образу жизни. Лигнин придает механическим волокнам и проводящим элементам растений, живущих на суше, необходимые жесткость и гидрофобность клеточных стенок. Благодаря лигнину и возникли на Земле древесные формы растений. В процессе образования лигнина одновременно возникают химические ковалентные и межмолекулярные, в том числе водородные, связи лигнина с углеводами - гемицеллюлозами, и таким образом строится лигнин-гемицеллюлозная матрица, в которой заключены микрофибриллы целлюлозы.
Лигнин чувствителен даже к сравнительно мягким обработкам. Поэтому он претерпевает значительные изменения при выделении.
2.4. Методы выделения и определение лигнина.
Выделение лигнина из древесины проводят с различными целями: для получения препаратов лигнина и их последующего исследования; для количественного определения лигнина в древесине и другом растительном сырье прямыми методами. При делигнификации сырья с целью получения технической целлюлозы и других волокнистых полуфабрикатов можно из отработанных варочных растворов выделить технические лигнины. В зависимости от цели подбирают соответствующие методы выделения. При получении препарата лигнина для исследования метод выделения должен обеспечить минимальное изменение самого лигнина. Выделить же природный лигнин из древесины в неизмененном состоянии практически невозможно. При количественном определении лигнина метод выделения должен обеспечить выход препарата лигнина, более или менее соответствующий его количеству в древесине. При делигнификации древесины в производстве целлюлозы основная задача заключается в получении целлюлозы с большим выходом и определенными показателями качества, в том числе с малым содержанием остаточного лигнина. В этом случае глубокие химические изменения, происходящие при его удалении, неизбежны. Технические лигнины, выделенные из отработанных варочных растворов, значительно изменены по сравнению с природным лигнином.
Для удаления экстрактивных веществ всегда необходимо предварительное экстрагирование, но любой растворитель вызывает потери лигнина. Чем жестче условия экстрагирования, тем больше потери лигнина.
Выделенный лигнин содержит примеси трех типов: примесь экстрактивных веществ, дающих в условиях кислотного гидролиза нерастворимые осадки; примесь продуктов гумификации сахаров — нерастворимых гуминоподобных соединений, образующихся из моносахаридов в жестких условиях кислотного гидролиза; примесь трудногидролизуемых полисахаридов и продуктов неполного гидролиза (олигосахаридов), адсорбируемых лигнином.
Наилучшими условиями гидролиза считают такие, в которых выделяется максимальное количество лигнина с максимальным содержанием метоксилов. Тогда выделенный лигнин не содержит примесей, ошибочно принимаемых за лигнин, и в то же время при его выделении не было значительных потерь.
Основное распространение в настоящее время получил сернокислотный метод определения лигнина. Солянокислотный метод приводит к меньшей гумификации Сахаров, но работа со сверхконцентрированной 41...42%-ной соляной кислотой менее удобна.
2.5. Получение препаратов лигнина.
При выделении лигнина из древесины с целью получения препаратов лигнина для исследования его строения, свойств и механизмов его реакций древесину предварительно измельчают до опилок или муки. Измельченную древесину освобождают от экстрактивных веществ (во избежание их возможной конденсации с лигнином) экстрагированием органическими растворителями, обычно спирто-толуольной смесью.
Вследствие существования химических связей лигнина с углеводами и сетчатой структуры самого лигнина для препаративного выделения лигнина необходимо применять кислотные катализаторы. Под действием кислотных катализаторов осуществляется сольволиз, например, гидролиз химических связей лигнина с гемицеллюлозами и частично связей в его сетчатой структуре с получением фрагментов сетки. Одновременно кислотные катализаторы в большей или меньшей степени способствуют конкурирующим реакциям конденсации, препятствующим переходу лигнина в растворимое состояние. Кроме того, реагенты и растворители, применяемые для получения препаратов лигнина, дают побочные реакции.
Все методы выделения лигнина можно подразделить на две группы: методы, основанные на удалении полисахаридов, с получением лигнина в виде нерастворимого остатка; методы, основанные на переводе лигнина в раствор с последующим осаждением (получение препаратов растворимых лигнинов). Нерастворимые препараты лигнина, имеющие подобно природному лигнину сетчатую структуру, более или менее сохраняют морфологическое строение клеточной стенки, но, естественно, более рыхлое. Растворимые лигнины после осаждения, очистки и сушки имеют вид порошков.
2.6 Получение кислотных лигнинов.
Эти методы основаны на гидролизе полисахаридов концентрированными минеральными кислотами. Различают сернокислотный лигнин (лигнин Класона), который получают обработкой древесины концентрированной (64...78%-й) серной кислотой и солянокислотный лигнин (лигнин Вильштеттера), который получают обработкой сверхконцентрированной (40...42%-й) соляной кислотой при охлаждении (температура 1... 5°С)
В лабораторной практике чаще используют серную кислоту. Гидролиз проводят в две ступени, сначала концентрированной кислотой при температуре около 20°С, а затем реакционную массу разбавляют водой и кипятят. Вторая ступень необходима, во-первых, для дополнительного гидролиза олигосахаридов, которые могут довольно прочно адсорбироваться лигнином, а, во-вторых, для осаждения из раствора лигнина, который частично растворяется (пептизируется) в концентрированной кислоте, особенно лигнин лиственной древесины. Гидролиз сверхконцентрированной соляной кислотой применяют реже из-за трудностей получения такой кислоты и работы с ней.
Препараты кислотных лигнинов имеют темный цве